Пантотенаткиназа
Бұл мақала әлі тексерістен өтпеді. Тексерілмеген мақалалардағы мәліметтер сенімсіз болуы мүмкін.
|
Пантотенаткиназа (EC 2.7.1.33, PanK; CoaA) коэнзим А (КоА) биосинтетикалық жолындағы бірінші фермент болып табылады. Ол аденозинтрифосфат(АТФ) молекуласы есебінен 4'-фосфопантотенат түзу үшін пантотенатты (В5 витамині) фосфорлайды. Бұл КоА биосинтезіндегі жылдамдықты шектейтін кезең.[1][2]
![[2]](http://upload.wikimedia.org/wikipedia/commons/3/34/Mechanism_os_pantothenate_kinase.png)
КоА барлық тірі организмдерге қажетті кофактор болып табылады. Ол лимон қышқылының циклі (үш карбон қышқылының циклі) және май қышқылының метаболизмі сияқты көптеген маңызды жасушалық процестерде негізгі ацил тобының тасымалдаушысы ретінде әрекет етеді. Демек, пантотенаткиназа CoA биосинтетикалық жолындағы негізгі реттеуші фермент болып табылады.[3]
Түрлері
өңдеуPanK-ның үш түрлі түрі анықталды - PanK-I (бактерияларда кездеседі), PanK-II (негізінен эукариоттарда, сонымен қатар стафилококктарда кездеседі) және PanK-III, сонымен қатар CoaX (бактерияларда кездеседі). Эукариоттық PanK-II ферменттері көбінесе PanK1, PanK2, PanK3 және PanK4 сияқты әртүрлі изоформалар түрінде кездеседі. Адамдарда бірнеше PanK изоформалары төрт генмен экспрессияланады. PANK1 гені PanK1α және PanK1β формаларын, ал PANK2 және PANK3 сәйкесінше PanK2 және PanK3 түрлерін кодтайды.[4]
Құрылым
өңдеуPanK-II
өңдеу
PanK-II құрамында 1-суретте көрсетілгендей екі ақуыз домені бар. A домені мен A' доменінің әрқайсысында глицинге бай цикл (GXXXXGKS тізбегі; P циклі) бар, ол нуклеотидтерді байланыстыру орындарына тән; бұл жерде ATP байланысады деп болжанады.[5] A доменіндегі 95 және 102 қалдықтары арасында орналасқан
Екі ATP байланыстыру алаңы бірлескен әрекетті көрсетеді . Димеризация интерфейсі бір-бірімен әрекеттесетін әрбір мономерден бір-бірден екі ұзын спиральдан тұрады. Спиральды ұштары әрбір мономердің валин мен метионин қалдықтары арасындағы ван-дер-Ваальс әрекеттесуімен бірге ұсталады. Спиральдардың ортасы аспарагин қалдықтары арасындағы сутектік байланыстармен бекітіледі. N-терминалдың соңында әрбір спираль кеңейіп, екі қысқа спиральдан тұратын төрт спираль шоғырын құрайды. Бұл байлам димерді одан әрі тұрақтандыру үшін ван-дер-Ваальс күштерін пайдаланатын полярлы емес қалдықтардан құралған гидрофобты өзектен тұрады.[4]
Белсенді аймақта пантотенат пантотенат пен аспартат, тирозин, гистидин, тирозин және аспарагин қалдықтарының бүйірлік тізбектері арасындағы сутектік байланыстармен бағдарланған.[6] Катализге аспарагин, гистидин және аргинин қалдықтары қатысады.
PanK1α, PanK1β, PanK2 және PanK3 адамның PanK-II изоформаларында шамамен 355 қалдықтың жалпы, жоғары гомологты каталитикалық өзегі бар.[4] PanK1α және PanK1β екеуі де PANK1 генімен кодталған және 2-ден 7-ге дейінгі экзондармен кодталған 363 аминқышқылдарының бірдей каталитикалық доменіне ие. PanK1α транскрипциясы N-терминуста 184 қалдық реттеуші доменді кодтайтын 1α экзонынан басталады. Бұл аймақ бос CoA және ацил-КоА арқылы кері байланысты тежеуге және ацетил-КоА және малонил-КоА арқылы реттеуге мүмкіндік береді. Екінші жағынан, PanK1β транскрипциясы кері байланысты реттеу доменін қамтымайтын 10 қалдық N-терминусын шығаратын экзон 1β-ден басталады.[7]
PanK-III
өңдеу
PanK-III құрамында екі ақуыздық домен бар және PanK-II негізгі каталитикалық қалдықтары сақталады. PanK-II және PanK-III мономер бірліктері іс жүзінде бірдей, бірақ олардың димерлі жинақтары анық ерекшеленеді. Staphylococcus aureus II типті және Pseudomonas aeruginosa III типті құрылымдар арасындағы зерттеу PanK-II мономерінде PanK-III мономерінде жоқ ілмек аймағы бар, ал PanK-III мономерінде жоқ цикл аймағы бар екенін көрсетеді. PanK-II мономерінен.[8] Бұл кішігірім вариацияның димеризация интерфейсінде шешуші айырмашылығы бар, онда PanK-II димер орамының спиралдары бір-бірінің айналасында және PanK-III димерінің спиралдары 70° бұрышта әрекеттеседі (2-сурет).[9]
PanK-II және PanK-III арасындағы димеризация интерфейсіндегі осы айырмашылықтың нәтижесінде АТФ пен пантотенат үшін субстратты байланыстыру орындарының конформациясы да ерекшеленеді. [10] [11]
Каталитикалық механизм
өңдеу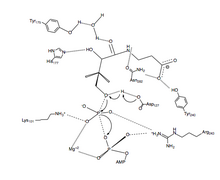
PanK-II
өңдеуPanK-II фосфорды тасымалдау реакциясының ұсынылған механизмі диссоциативті ауысу күйі бар келісілген механизм болып табылады.
Біріншіден, АТФ P контурының қалдықтары және жақын орналасқан қалдықтар арқылы жасалған байланыстырушы ойықта байланысады. Мұнда сақталған лизин (Lys-101) ATP байланысуы үшін қажетті негізгі қалдық болып табылады.[12][13] Сонымен қатар, Lys-101, Ser-102, Glu-199 және Arg-243 қалдықтарының бүйірлік тізбектері байланыстырушы ойықтағы нуклеотидті бағыттайды. Пантотенат Asp-127, Tyr-240, Asn-282, Tyr-175 және His-177 қалдықтарымен сутектік байланыс түзу арқылы байланысқан және бағытталған.[6] ATP және пантотенат екеуі де байланысқан кезде, Asp-127 пантотенаттың C1 гидроксил тобын протонациялайды. Содан кейін пантотенаттан шыққан оттегі байланысқан АТФ-ның γ-фосфатына әсер етеді. Мұнда β-және γ-фосфат топтарының зарядының тұрақтануына Arg-243, Lys-101 және үйлестірілген Mg2+ иондары қол жеткізеді.[14] Бұл келісілген механизмде γ-фосфаттың жазық фосфораны пантотенаттың шабуылдаушы оттегіне желіде тасымалданады. [6] Соңында, 4'-фосфопантотенат PanK-дан диссоциацияланады, содан кейін ADP.
Пантотенаткиназаның реттелуі
өңдеуPanK-II
өңдеуПантотенаткиназаның реттелуі жасушаішілік CoA концентрациясын бақылау үшін маңызды.[15] Пантотенаткиназа КоА және оның тиоэфирлері (яғни, ацетил-КоА, малонил-КоА ) арқылы кері байланыс тежеу арқылы реттеледі.[16] КоА ATP байланыстыру алаңымен бәсекелесу және Lys-101-ге ATP байланысуын болдырмау арқылы PanK белсенділігін тежейді.[12][13]
КоА АТФ -мен бір жерде байланысқанымен, олар әртүрлі бағытта байланысады және олардың аденин бөліктері ферментпен қабаттаспайтын қалдық жиынтығымен әрекеттеседі. His-177, Phe-247 және Arg-106 CoA тану үшін қажет, бірақ ATP үшін емес, және Asn-43 және His-307 АТФ адениндік негізімен әрекеттессе, His-177 және Phe-247 аденинмен әрекеттеседі. CoA негізі.[14] Екі молекула да өздерінің фосфоэфирлеріндегі зарядты бейтараптандыру үшін Lys-101 пайдаланады.
Эфирленбеген КоА тиоэфирлеріне қарағанда күшті тежеу қабілетіне ие. Бұл құбылыс тиол тобының Phe-244, Phe-259, Tyr-262 және Phe-252 хош иісті қалдықтарымен тығыз орналасуымен жақсы түсіндіріледі. Бос CoA оңтайлы сәйкестікке ие, бірақ ацил тобы CoA-ға қосылған кезде, стерикалық кедергі тиоэфирдің Phe-252-мен әрекеттесуін қиындатады. Осылайша, тиоэфирлермен тежелу эфирленбеген КоА-ға қарағанда тиімдірек.[14]
PanK-III
өңдеуЖоғарыда сипатталған ереже PanK-II сәйкес келеді. PanK-III кері байланыс тежелуге төзімді.[8][10] [11]
Гендер
өңдеуАдамдарда:
- PANK1, PANK2, PANK3, PANK4.
PANK2 гені жасушаның энергия өндіретін орталықтары болып табылатын митохондрияларда КоА түзілуін реттейтін PanK2-ні кодтайды.[17] PANK2 мутациясы бұрын Халлерворден-Спатц синдромы деп аталатын пантотенаткиназамен байланысты нейродегенерацияның себебі болып табылады. Бұл сирек ауру терең дистониямен, спастикалықпен көрінеді және жиі өліммен аяқталады.
Дереккөздер
өңдеу- ↑ "Rate-limiting step and control of coenzyme A synthesis in cardiac muscle". The Journal of Biological Chemistry 257 (18): 10967–72. September 1982. PMID 7107640.
- ↑ a b "Crystal structure of a type III pantothenate kinase: insight into the mechanism of an essential coenzyme A biosynthetic enzyme universally distributed in bacteria". Journal of Bacteriology 188 (15): 5532–40. August 2006. PMID 16855243.
- ↑ "Coenzyme A: back in action". Progress in Lipid Research 44 (2–3): 125–53. 2005-03-01. doi:10.1016/j.plipres.2005.04.001. PMID 15893380.
- ↑ a b c "Crystal structures of human pantothenate kinases. Insights into allosteric regulation and mutations linked to a neurodegeneration disorder". The Journal of Biological Chemistry 282 (38): 27984–93. September 2007. PMID 17631502.
- ↑ "The P-loop--a common motif in ATP- and GTP-binding proteins". Trends in Biochemical Sciences 15 (11): 430–4. November 1990. doi:10.1016/0968-0004(90)90281-F. PMID 2126155.
- ↑ a b c "The structure of the pantothenate kinase.ADP.pantothenate ternary complex reveals the relationship between the binding sites for substrate, allosteric regulator, and antimetabolites". The Journal of Biological Chemistry 279 (34): 35622–9. August 2004. PMID 15136582.
- ↑ "The murine pantothenate kinase (Pank1) gene encodes two differentially regulated pantothenate kinase isozymes". Gene 291 (1–2): 35–43. PMID 12095677.
- ↑ a b "Structural basis for substrate binding and the catalytic mechanism of type III pantothenate kinase". Biochemistry 47 (5): 1369–80. February 2008. doi:10.1021/bi7018578. PMID 18186650.
- ↑ "Prokaryotic type II and type III pantothenate kinases: The same monomer fold creates dimers with distinct catalytic properties". Structure 14 (8): 1251–61. August 2006. doi:10.1016/j.str.2006.06.008. PMID 16905099.
- ↑ a b "Characterization of a new pantothenate kinase isoform from Helicobacter pylori". The Journal of Biological Chemistry 280 (21): 20185–8. May 2005. doi:10.1074/jbc.C500044200. PMID 15795230.
- ↑ a b "Inhibitors of pantothenate kinase: novel antibiotics for staphylococcal infections". Antimicrobial Agents and Chemotherapy 47 (6): 2051–5. June 2003. doi:10.1128/AAC.47.6.2051-2055.2003. PMID 12760898.
- ↑ a b "Kinetics and regulation of pantothenate kinase from Escherichia coli". The Journal of Biological Chemistry 269 (43): 27051–8. October 1994. PMID 7929447.
- ↑ a b "Cloning, sequencing, and expression of the pantothenate kinase (coaA) gene of Escherichia coli". Journal of Bacteriology 174 (20): 6411–7. October 1992. PMID 1328157.
- ↑ a b c "Structural basis for the feedback regulation of Escherichia coli pantothenate kinase by coenzyme A". The Journal of Biological Chemistry 275 (36): 28093–9. September 2000. PMID 10862768.
- ↑ "Regulation of coenzyme A biosynthesis". Journal of Bacteriology 148 (3): 926–32. December 1981. PMID 6796563.
- ↑ "Role of feedback regulation of pantothenate kinase (CoaA) in control of coenzyme A levels in Escherichia coli". Journal of Bacteriology 185 (11): 3410–5. June 2003. doi:10.1128/JB.185.11.3410-3415.2003. PMID 12754240.
- ↑ PANK2 gene. Genetics Home Reference (22 ақпан 2016). Тексерілді, 29 ақпан 2016.
Сыртқы сілтемелер
өңдеу- Pantothenate+kinase at the US National Library of Medicine Medical Subject Headings (MeSH)
- EC 2.7.1.33